Image credit Natural History Museum-London
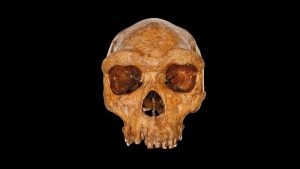
Fossils assigned to Homo heidelbergensis (named for a mandible found near Heidelberg, Germany) are found throughout the Old World from tropical to temperate zones at sites dating to the Middle Pleistocene between 800,000 and 125,000 years ago. Some of these sites include Bodo and Kabwe in Africa; Petralona, Arago, and Sima de los Huesos in Europe; and Dali and Jinnishuan in Asia. Homo heidelbergensis displays traits that are primitive (traits shared with its ancestor, in this case, Homo erectus); however, it also possesses many derived traits (traits different from those found in the ancestral species, in this case, traits that are more similar to those found in Homo sapiens). There is evidence that H. heidelbergensis made fairly sophisticated stone tools and hunted large animals, suggesting an advanced ability to engage in cooperative social activities. Because the size of the sample of H. heidelbergensis fossils is small and many fossils have not been precisely dated, the relationships between this species and those that came before and after (as well as the cohesiveness of the species itself) is the source of substantial debate among scientists. Paleoanthropologists often refer to the uncertainties surrounding the specimens, their dating and morphology, as “the muddle in the middle” (referring to the “Middle” Pleistocene).
It is difficult to create a list of features that definitively distinguish H. heidelbergensis from its likely predecessor (H. erectus) and its purported descendants Homo neanderthalensis and H. sapiens. Features that link H. heidelbergensis to H. erectus include a massive, projecting face and brow ridge (the bony ridge over the eye, also known as the supraorbital torus), a low frontal bone (a forehead that does not extend greatly above the level of the brow ridges), and sagittal keeling, a thickening of bone along the top of the skull from front to back. Homo heidelbergensis also possesses derived characteristics aligning it with H. sapiens. For instance, fossils attributed to this species exhibit separate supraorbital tori (brow ridges) over each orbit (unlike the continuous, single brow ridge found in H. erectus), a posterior cranial vault (the back of the skull) that is more vertical than in H. erectus, parietal bones (bones that form part of the sides and top of the skull) that are wide relative to the cranial base (underside of skull) compared to H. erectus, and a large cranial capacity (volume of part of the skull that approximates brain size) in the approximate range of H. sapiens.
Some of the earliest H. heidelbergensis fossils, dating roughly between 600,000 and 400,000 years ago, are found in Africa. Kabwe, one of the most famous H. heidelbergensis specimens, was found at a mining site in 1921 and cannot be accurately dated, but most researchers agree that it is Middle Pleistocene in age (at least 125,000 years ago and possibly earlier). Other sites in Africa, including Bodo (dated to about 600,000 years ago, now considered by some scientists the type specimen of a new species, Homo bodoensis), contain stone tools resembling the Acheulean tools used by H. erectus. However, the tools used by H. heidelbergensis were thinner and more finely flaked than those used by H. erectus, leading researchers to hypothesize the change in tool manufacture reflects a behavioral change that can be linked to a speciation event (an evolutionary process by which new species arise) in which H. erectus gave rise to a new species with a larger brain and possessing the capability of engaging in more complex behaviors. Along with stone tools, the remains of hippos, baboons, and antelopes with cut marks (marks made by stone tools) on the bone are often found at Middle Pleistocene sites in Africa, suggesting that H. heidelbergensis was butchering meat. Cut marks on the face around the eye sockets of the Bodo cranium have suggested it was also butchered.
Much of the information we have about H. heidelbergensis comes from sites in Europe. Some fossil skulls from the Middle Pleistocene in Europe very closely resemble those found in Africa. Examples are a skull from Petralona, Greece (whose exact date is unknown but is likely Middle Pleistocene in age), and a partially distorted facial skeleton from Arago in France (dating roughly to 450,000 years ago). Some scientists recognize features in European H. heidelbergensis that seem to foreshadow features distinctive of H. neanderthalensis. For example, the face of the Arago fossil, according to some researchers, has a flattened cheek where the zygomatic arches (cheek bones) appear “swept back” as in Neandertals. However, not all researchers agree that this is the case, especially since the Arago face is distorted. Two lower jaws have also been found at Arago. These closely resemble the Mauer mandible (the type specimen or fossil which serves as the basis on which H. heidelbergensis was named, see above) and in so doing, corroborate the attribution of the Arago fossil to H. heidelbergensis. This similarity also underlines the general morphological similarities among the African and European fossils attributed to Homo heidelbergensis.
In Spain, the site of Sima de los Huesos (400,000 years ago) has yielded a spectacular amount of postcranial (the skeleton minus the skull) remains along with a complete skull. The postcranial remains show that the individuals from Sima de los Huesos were tall, robust, and had modern levels of sexual dimorphism (difference in size between the sexes). Researchers have suggested the fossils found at Sima de los Huesos have features connecting them to H. neanderthalensis; for example, the infraorbital surface (the face below the eye socket) and side wall of nose meet at shallow angle, producing a concavity that may foreshadow H. neanderthalensis morphology. Furthermore, the brows are robust, double-arched, and continuous across the forehead at the glabella (the area of bone between the eyebrows), and the nasal bones are broad, which are also characteristics commonly associated with H. neanderthalensis.
However, whether these traits are true H. neanderthalensis apomorphies (a trait unique to a particular species and all its descendants) is unclear. The relationship between European H. heidelbergensis and populations outside of Europe differs depending on how these traits are interpreted.
Fossils found at Gran Dolina (780,000 years ago) in Spain contribute to the debate over the relationship between European and African Middle Pleistocene hominins. A hollow cheek and vertical nasal aperture combined with a mandibular joint resembling that of Homo sapiens underline the distinctiveness of these fossils vis-à-vis H. erectus. They also possess no specialized Neandertal traits but have teeth that resemble other African and European Middle Pleistocene hominins. Furthermore, at least one fossil has a canine fossa (a depression in the facial skeleton to the side of the canine tooth socket), a trait usually associated with Homo sapiens. Based on this mixture of traits, some scientists have placed the Gran Dolina fossils into a new species, “Homo antecessor.” However, the sample for this species is small, fragmentary, and mostly subadult (non-adult specimens). Furthermore, a canine fossa is variably expressed in other Middle Pleistocene hominin populations (like those in China, see below), so the significance of this feature is unclear. Overall, only a few traits differentiate the Gran Dolina fossils from other European fossils in the Middle Pleistocene, and, at present, the evidence suggesting the Gran Dolina specimens represent a distinct species is scant. Many researchers attribute these fossils to H. heidelbergensis.
Asian sites have yielded Middle Pleistocene fossils with variable characteristics that some scientists believe link these fossils to African and European H. heidelbergensis. There is no agreement, however, regarding whether these fossils can be easily so attributed. The Narmada fossil, from India, possesses morphology generally similar to African and European H. heidelbergensis. While this fossil is securely dated to the Middle Pleistocene, the precise date is not known. In China, two Middle Pleistocene sites, Dali and Jinnishuan (both dated to between 300,000 and 200,000 years ago) have yielded crania resembling H. heidelbergensis from Africa and Europe.
The morphology of H. heidelbergensis is a mosaic of ancestral and derived traits, fueling a debate over which fossils should be included within the species and whether the species itself is valid. Traditionally, H. heidelbergensis specimens have been lumped together in a loosely defined species called “archaic Homo sapiens,” a group that includes fossils that are too modern to be H. erectus but not modern enough to fit comfortably within H. sapiens. Archaic H. sapiens has been described as a “wastebasket taxon,” into which Middle Pleistocene fossils are placed when it is not clear to which species they belong. The debate over Middle Pleistocene taxonomy stems from the fact that dating is generally poor for sites from this period, the fossil sample is relatively small, and different scientists interpret morphologies in different ways.
It has been determined that by 1.6 million years ago, H. erectus was well-established outside of Africa in both Europe and Asia. Around 780,000 years ago, hominins begin to appear with morphology that was different from H. erectus in southern Europe and Africa. The phylogeny of hominins in the Middle Pleistocene depends on how certain characteristics are interpreted. For example, some scientists agree that European and African Middle Pleistocene hominins shared a robust body form as well as derived features tied to expansion of brain and cranial vault. Interpreting Middle Pleistocene hominin morphology in this way leads to the conclusion that all early Middle Pleistocene fossils belong to the same lineage in a single species called “Homo heidelbergensis.” Later, some populations moved north to Europe where cold adaptation eventually led to the evolution of H. neanderthalensis. Simultaneously, H. heidelbergensis populations in Africa evolved into modern humans.
Some scientists prefer a different interpretation, according to which H. antecessor is descended from H. erectus in Africa and gives rise to all later humans. In this scenario, H. antecessor evolved into an exclusively European H. heidelbergensis in Spain, which gradually evolved into H. neanderthalensis via anagenesis (e.g., evolution within a species without the origination of a new species). According to this view, H. antecessor gave rise to Homo rhodesiensis in Africa, a species that includes specimens like Kabwe and Bodo, which later evolved into Homo sapiens in another anagenetic scenario. Dating of the Asian Middle Pleistocene fossils is particularly problematic, but it is probable that many of the Asian fossils are geologically younger than European and African H. heidelbergensis. These Asian fossils could perceivably represent an eastward migration of H. heidelbergensis late in the Middle Pleistocene. However, evidence for this link is tenuous at best, and firm dating along with more fossil evidence is needed to test this hypothesis.
One of the most important facts about H. heidelbergensis is the archaeological record (artifacts, including tools, and remains of butchered animals) suggests this species had cognitive abilities that were advanced relative to those possessed by H. erectus. In Africa, Middle Pleistocene sites yield stone tools that are reminiscent of H. erectus Acheulean technology, but with notable differences. For instance, the Middle Pleistocene (so called “Later” or “Developed Acheulean”) tools are thinner, more symmetrical, and have more flake scars (evidence of flakes of stone that were chipped off the tool in the course of its manufacture) than H. erectus tools. At some sites, small hand axes are accompanied by flake tools (tools formed from the flakes chipped off the source rock, in contrast to “core tools,” which are tools formed by chipping off flakes from the source rock) like those of Middle Stone Age in Africa (roughly dated to 300,000 to 50,000 years ago).
Several archaeological finds from Europe also support the interpretation of increased behavioral sophistication in H. heidelbergensis. The site of Boxgrove in England (dated to roughly 500,000 years ago) has yielded thin, extensively flaked flint bifaces (a stone tool that is produced by removing flakes of stone from both sides) along with cut-marked bones of horses and rhinos. A large, robust hominin tibia (i.e., shin bone), attributed by some to H. heidelbergensis, has also been recovered at Boxgrove. In Germany, the site of Schöningen (dated to roughly 400,000 years ago) has yielded three wooden spears associated with flint tools and chips. Similar sites in Spain, France, and Italy also yield Late Acheulean tools associated with large herbivore bones. Together, these finds are evidence that H. heidelbergensis may have hunted and butchered large game, an activity that requires experience, practice, and cooperation. Based on archaeological evidence, H. heidelbergensis had increased levels of social cooperation.