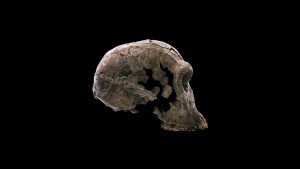
Homo habilis, which dates to between 2.3 and 1.5 million years ago is named for the Latin term “habilis,” meaning “handy, skillful, able” and is one of the earliest species in the genus Homo. Fossils of H. habilis have been found in Tanzania, Ethiopia, Kenya, and South Africa and represent cranial (from the skull), dental (teeth), and postcranial (from the skeleton below the skull) remains. While this species is distinct from the australopiths in many aspects of its cranial morphology (size and shape), it also exhibits many primitive traits (shared with its ancestor, in this case species in the genus Australopithecus), especially in its postcranial skeleton, which suggest it was more similar to its australopith ancestors than was originally thought.
Compared to australopiths, H. habilis has a relatively and absolutely bigger brain (average cranial capacity—a measure of the volume of the braincase, used to estimate brain volume—is around 680 cubic centimeters), a more vertical forehead, and weaker brow ridges (the enlarged bony area over the eye sockets). Additionally, the face and jaws of H. habilis are smaller and less projecting than those of the australopiths, and the cheek teeth (the teeth behind the canine—molars and premolars) are narrower mesiodistally (front-to-back).
Postcranial remains of H. habilis indicate a mosaic of primitive and derived traits (traits different from those in the ancestral species, in this case different from the australopiths and more similar to later species in the genus Homo). For example, H. habilis was undoubtedly bipedal, with an adducted big toe (the big toe is in line with the other toes, not splayed out to the side, as in living apes) and well-defined arches in the foot (structures in the foot formed by bone and strengthened by tendons and ligaments that allow the foot to support the weight of the body and act as shock-absorbers during bipedal walking) like humans. However, while H. habilis shows definite signs of bipedalism, it also shared features with great apes, such as a marked tubercule (enlargement of bone at a site of muscle attachment) for a leg muscle useful for climbing. While only a few postcranial remains associated with H. habilis permit reconstructions of the proportions of the limbs, the fossils that have been found suggest that the legs are elongated relative to australopiths but have retained long forearms. The H. habilis hand is a mosaic of ape- and human-like features. The phalanges (finger bones) resemble those of living apes insofar as they are robust (thick) and curved but have broad tips (similar to humans) and attach to the palms in a way similar to humans. Additionally, some bones of the wrist and attachment sites for flexor tendons (tendons that pull the palm of the hand closer to the forearm) are more ape-like and have been suggested to be useful while climbing.
Because of the range of variation present in early Homo fossils, there has been controversy over whether H. habilis should be considered one highly variable, sexually dimorphic (exhibiting large size differences between the sexes) species or whether some specimens originally grouped with H. habilis should be separated into a different species, Homo rudolfensis (see essay on H. rudolfensis for more detailed information on this controversy). However, critical examination of the fossil anatomy of these specimens reveals that the patterns of variation expected in a sexually dimorphic species are not present among H. habilis and H. rudolfensis fossils. Furthermore, dental analysis suggests that each species had very different ecological niches. At present, it is generally accepted that H. habilis and H. rudolfensis represent separate species.
The first H. habilis fossils were found in Olduvai Gorge in the same stratigraphic layer as early types of stone tools called Oldowan tools. These tools are very simple and comprise a core—the scarred center of rock that results from the detachment of one or more flakes (the portion of rock removed from original source material by percussion or pressure)—from a lump of source material. These cores, while simple, had sharp, cutting edges and could be used for many purposes. Although H. habilis fossils are found near stone tools, they are not the only hominins to be linked to tool manufacture. For example, Paranthropus boisei fossils have also been found near stone tools, making definitive determination of which species was a toolmaker impossible. (There were several different species of Australopithecus, Paranthropus, and Homo, alive between 2.5 and 2.0 million years ago, as can be seen on the Timeline). Because H. habilis has a larger brain and smaller teeth than other australopiths (including P. boisei), many scientists consider them to be the earliest toolmakers. However, this assertion may need to be reevaluated in light of more recent finds of a species of australopith, Australopithecus garhi, with a smaller brain and larger teeth than H. habilis, and that is associated with stone tools (see essay on A. garhi for more information). Speculations as to whether Australopithecus or Homo was the first user of stone tools may have been made moot by the discovery in 2009 of markings on animal bones that could have been made only by stone being used to deflesh the bones. The bones bearing stone cut marks were found at Dikika, Ethiopia, and dated to 3.4 million years, a time at which the only hominin then extant was Australopithecus afarensis.
About 2.5 million years ago, Africa underwent a climatic change that resulted in the changing of hominin habitats from more closed and wet to more open and arid, which led to changes in available food resources and may have signaled the origins of the genera Homo and Paranthropus. By approximately 2.0 million years ago, however, the climate was returning to slightly more warm and humid conditions. Environmental reconstructions of H. habilis sites suggest that these hominins lived in a woodland habitat with access to lakes and/or streams.
H. habilis is one of the oldest species in the genus Homo. Nevertheless, evidence suggests that in some ways, it was quite similar to species in the genus Australopithecus, especially in aspects of the postcranial skeleton and the small size of its brain. Taking into account body size and shape, locomotion, the masticatory system, and brain size, some scientists suggest that H. habilis had an adaptive strategy more similar to australopiths than to modern humans and should be placed within the genus Australopithecus. Whether or not this is a valid suggestion depends upon how a genus is defined. Scientists disagree as to whether phylogeny (evolutionary relationships) should be given priority over adaptive strategies when defining a genus, or vice versa, a distinction that is not easy to make, especially when dealing with fossil specimens. Currently, H. habilis is placed within the genus Homo because it shares derived traits with other members of the genus to the exclusion of the australopiths.
Assigning H. habilis to either the genus Homo or Australopithecus has implications for the way we interpret the fossil record. If H. habilis belongs in the genus Homo, this suggests that the beginning of our evolutionary line encompassed a very broad range of morphological and adaptive variation. The variation present at the beginning of our clade (a group consisting of an ancestor and all of its descendants) suggests that human evolutionary history is more complicated than has traditionally been assumed.