Image credit National Geographic, courtesy Youseuke Kaifu; University of Wollongong
Homo floresiensis is a small-bodied hominin species that is dated to between 100,000 and 50,000 years ago. Known from a single cave on the island of Flores in eastern Indonesia, H. floresiensis displays a surprising combination of cranial features that resemble those of Homo erectus and Homo sapiens, postcranial features (features of the limb and trunk) that most closely resemble those of species in the genus Australopithecus, and a stone tool technology linked to those often associated with H. sapiens. If the fossils assigned this species do not represent a population of pathological H. sapiens, as some researchers have argued, they represent a new hominin species and suggest H. sapiens shared the planet with another species much more recently than previously believed.
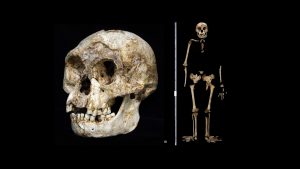
The cranial material of H. floresiensis comprise teeth and mandibles (lower jaws) from multiple individuals and a single, nearly complete skull. The skull and teeth share some derived features (features not found in the common ancestor) shared with H. erectus and H. sapiens. The face is small, specifically in facial height (a measurement between the brow and the upper teeth) and exhibits reduced prognathism (how far the upper and lower jaws project out from the face) compared to australopith species. The H. floresiensis molars (the teeth at the back of the jaw) and canines are also relatively small, similar to the condition found in H. erectus and H. sapiens. These Indonesian fossils also possesses many primitive features similar to earlier hominins. For example, the brain is very small (380 to 420 cubic centimeters) and is more similar to apes and species in the genus Australopithecus. The mandibles and premolars (bicuspid teeth), which represent at least eight partial individuals, share distinctive primitive traits with both Australopithecus and Homo habilis. The premolars are asymmetrical and elongated with complex tooth roots. The mandibles are relatively robust and large in size, yet lack a chin (central part of the lower jaw that protrudes out from the face, found only in H. sapiens mandibles).
In contrast to the cranium, where only one complete skull was found, postcranial remains from multiple individuals have been recovered. The shoulder joint is cranially oriented (toward the skull) and the clavicle (collar bone) is short and very curved, suggesting the shoulder was high on the torso, as seen in apes and species in the genus Australopithecus. Other aspects of the postcranial skeleton of these Flores finds that are shared with australopiths include relatively long arms, flared pelves (plural of pelvis; pelvic blades that extend beyond the hip joint) and relatively short legs. Moreover, the hand and wrist of H. floresiensis seem to be more primitive than even the earliest representatives of the genus Homo (H. habilis) and closely resemble the shape found in australopiths. The primitive morphology (size and shape) of the H. floresiensis hand and wrist suggests this species lacked the ability to precisely manipulate items with their hands. Interestingly, the foot of this species is similar to H. sapiens in some respects: the big toe is aligned with the other toes, and the bones are thick and robust. Nevertheless, the overall foot morphology exhibits a generally primitive pattern. For instance, the foot is approximately 20 centimeters in length, much longer than any H. sapiens foot from an individual of comparable height to the short statured H. floresiensis individuals, resembling more closely the length of chimpanzee and australopith foot. The toes exhibit primitive features such as long and curved lateral toes (the four toes to the outside of the feet, all but the hallux or big toe), and the hallux is quite short in comparison to that possessed by H. sapiens. The H. floresiensis foot also seems to lack the longitudinal arch that is characteristic of H. erectus and H. sapiens.
Some researchers argue the Flores remains are those of a diseased individual of our own species and have suggested a number of pathologies that could explain the mosaic of features exhibited by these remains. Pathologies that have been suggested include various forms of microcephaly (a class of genetic disorders in which the head and brain are abnormally small) and growth disorders, such as hypothyroidism (a condition in which the thyroid gland does not produce enough hormones that regulate the body’s metabolism) and Laron’s Syndrome (a genetic disorder in which the body does not process growth hormones normally). Detailed comparisons of the H. floresiensis material, however, seem to provide little support for these hypotheses because measurements of the remains fall outside of modern H. sapiens ranges of variation, even when compared to those with the aforementioned pathological conditions. Moreover, no pathological syndrome seems to adequately explain the suite of features exhibited by H. floresiensis. The fact that all of the individuals found at the cave site exhibit similar cranial and postcranial morphology casts further doubt on the hypothesis that the H. floresiensis remains are those of pathological H. sapiens, as it is unlikely multiple individuals would show signs of relatively rare diseases.
Considerable attention has also been paid to the evolutionary history of H. floresiensis—e.g., how did it get to Indonesia and of which previously known hominin species is it a descendant? Some researchers have suggested H. floresiensis represents a dwarfed descendent of Asian H. erectus populations. However, there is no evidence for larger-bodied (non-dwarfed) ancestors of H. floresiensis on the island, making this hypothesis difficult to test. This fact and H. floresiensis’ primitive anatomy (brain size, limbs, pelvis, hands, and wrist) has led some anthropologists to postulate the remains are evidence of a pre-H. erectus migration of an earlier species in the genus Homo or a species in the genus Australopithecus. Another hypothesis is that H. floresiensis descended from an earlier, more primitive type of H. erectus (such as that represented by H. erectus remains from the site of Dmanisi in the Republic of Georgia; see essay on H. erectus). Most researchers agree the current anatomical evidence shows that H. floresiensis is in many ways more similar to early Homo species (e.g., Homo habilis) than to later Homo species. This observation supports the idea that the ancestors of H. floresiensis left the African continent before the evolution of H. erectus, but the precise origins of this species remain unknown.
The stone artifact assemblages found in association with the skeletal remains of H. floresiensis are dense and demonstrate continuity in production method and tool type throughout the cave deposits. There are a large number of bifacial (struck on both sides) and radial (struck from the outer edge towards the center) cores (rocks from which pieces [flakes] are chipped to produce tools), similar to the Oldowan stone artifact assemblages in East Africa. More complex tools—e.g., points (sharpened, pointed tools), perforators (tools designed to make holes or incisions), blades (flake that is at least twice as long as it is wide), and microblades (blades less than 10 mm, often components of composite tool technology)—have also been recovered. The combination of primitive, Oldowan-like core technology with more advanced tool assemblages is quite uncommon in the archaeological record and make interpretations of the stone tool assemblages found in association with H. floresiensis difficult. Some researchers have questioned whether H. floresiensis had the cognitive capacity to make the more complex tools, citing its small brain size. However, no skeletal remains of H. sapiens have been found at sites bearing H. floresiensis skeletal material and artifacts commonly associated with the symbolic behaviors of H. sapiens (e.g., personal ornaments, pigments, and formal disposal of the dead) are also absent, suggesting that H. floresiensis was the manufacturer of all the tools found at this site.
Researchers have recently recovered stone tools from a site close to the cave on Flores Island. This site dates to roughly 1.02 million years ago and therefore provides a new minimum age for hominin inhabitation of the island. The artifacts found at this site are similar to those found at the original site; they are small flakes and both bifacially and radially struck cores. However, the absence of skeletal remains at this second site disallows the establishment of a direct link between H. floresiensis (or any other hominin species) and this stone tool assemblage.
The reconstructed paleoenvironment for H. floresiensis spans three cycles of glacial (cold, dry periods) to interglacial (warm episodes between glacial intervals) changeovers. The earlier cave sediments suggest a wet, rich surrounding environment with dense forests. However, a shift is indicated after 39,000 years ago, and Flores begins a transition to more arid conditions. There is a reduction in rainfall and forest cover as well as the development of widespread grasslands. The environment changed yet again 17,000 years ago and became gradually more wet. The appearance of parrots in the assemblage suggests that the local flora was changing into a more closed canopy forest, similar to present day environments.